Erwin Schrödinger - ¿Qué es la Vida?
Aquí puedes leer online Erwin Schrödinger - ¿Qué es la Vida? texto completo del libro (historia completa) en español de forma gratuita. Descargue pdf y epub, obtenga significado, portada y reseñas sobre este libro electrónico. Año: 1944, Editor: ePubLibre, Género: Ordenador. Descripción de la obra, (prefacio), así como las revisiones están disponibles. La mejor biblioteca de literatura LitFox.es creado para los amantes de la buena lectura y ofrece una amplia selección de géneros:
Novela romántica
Ciencia ficción
Aventura
Detective
Ciencia
Historia
Hogar y familia
Prosa
Arte
Política
Ordenador
No ficción
Religión
Negocios
Niños
Elija una categoría favorita y encuentre realmente lee libros que valgan la pena. Disfrute de la inmersión en el mundo de la imaginación, sienta las emociones de los personajes o aprenda algo nuevo para usted, haga un descubrimiento fascinante.

- Libro:¿Qué es la Vida?
- Autor:
- Editor:ePubLibre
- Genre:
- Año:1944
- Índice:5 / 5
- Favoritos:Añadir a favoritos
- Tu marca:
¿Qué es la Vida?: resumen, descripción y anotación
Ofrecemos leer una anotación, descripción, resumen o prefacio (depende de lo que el autor del libro "¿Qué es la Vida?" escribió él mismo). Si no ha encontrado la información necesaria sobre el libro — escribe en los comentarios, intentaremos encontrarlo.
Erwin Schrödinger: otros libros del autor
¿Quién escribió ¿Qué es la Vida?? Averigüe el apellido, el nombre del autor del libro y una lista de todas las obras del autor por series.












¿Qué es la Vida? — leer online gratis el libro completo
A continuación se muestra el texto del libro, dividido por páginas. Sistema guardar el lugar de la última página leída, le permite leer cómodamente el libro" ¿Qué es la Vida? " online de forma gratuita, sin tener que buscar de nuevo cada vez donde lo dejaste. Poner un marcador, y puede ir a la página donde terminó de leer en cualquier momento.
Tamaño de fuente:
Intervalo:
Marcador:
CAPÍTULO 3
Und was in shwankender Erscheinung schwebt, Befestiget mit dauernden Gedanker.
Goethe
3.1. Mutaciones discontinuas: material de trabajo de la selección natural
Los hechos generales que acabamos de exponer sobre la duración atribuida a la estructura del gen son quizá demasiado familiares como para sorprendernos o incluso para considerarlos convincentes. Esta vez, al menos, el refrán, según el cual la excepción confirma la regla, se convierte en realidad. Si el parecido entre padres e hijos no presentara excepciones, no solo no dispondríamos de todos esos brillantes experimentos que nos han revelado el detallado mecanismo de la herencia, sino que no se daría ese experimento a gran escala de la Naturaleza, realizado algunos millones de veces ya, que forja las especies por selección natural y supervivencia del más apto.
Tomemos este importante punto como principio para presentar los hechos principales, de nuevo pidiendo excusas y recordando que no soy biólogo.
Hoy sabemos con certeza que Darwin estaba en un error al considerar las variaciones pequeñas y continuas que se presentan incluso en la población más homogénea como los fenómenos sobre los que actúa la selección natural. Se ha demostrado que esas variaciones no se heredan. El hecho es suficientemente importante como para ilustrarlo con brevedad. Si tomamos la recolección de una raza pura de cebada, medimos una por una la longitud de las espigas y construimos una gráfica con los resultados, obtendremos una curva acampanada como la de la figura 1, donde en ordenadas se indica el número de espigas de una determinada longitud y en abscisas las diversas longitudes de espiga posible. En otras palabras: domina una longitud media definida, existiendo desviaciones con determinadas frecuencias en cada dirección. Elijamos un grupo de espigas (indicado en negro en la figura) con longitudes considerablemente mayores que la media, pero cuyo numero es suficiente para que puedan ser sembradas y den una nueva cosecha. Al hacer la estadística correspondiente, Darwin habría esperado encontrar que la curva se hubiese desplazado hacia la derecha. En otras palabras, se habría esperado producir por selección un aumento de la longitud media de las espigas. Esto no ocurre si se ha empleado una raza de cebada verdaderamente pura. La nueva curva estadística obtenida de la cosecha seleccionada es idéntica a la primera, y habría ocurrido lo mismo en caso de seleccionar para la siembra las espigas especialmente cortas. La selección resulta ineficaz porque las diferencias pequeñas y continuas no se heredan. Es obvio que no radican en la estructura del material hereditario, sino que, evidentemente, son accidentales. No obstante, el holandés Hugo de Vries descubrió hace unos cuarenta años que incluso en la descendencia de cepas realmente puras un número muy pequeño de individuos, algo así como dos o tres entre varias decenas de miles, aparece con cambios pequeños, pero que suponen una especie de «salto».
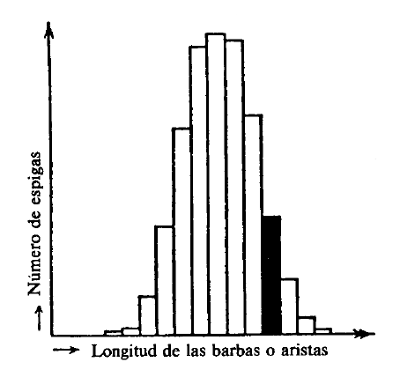
F IGURA 1. Gráfico estadístico de la longitud de las barbas de las espigas de una raza pura de cebada. Seleccionaremos para la siembra de semillas, del grupo indicado en negro. (Los datos del grafico no se refieren a un experimento real, pero sirven de ejemplo).
La expresión «salto» no quiere significar que el cambio sea especialmente importante, sino que supone una discontinuidad, en el sentido de que no hay formas intermedias entre la forma inalterada y los pocos individuos que han cambiado. Tras su observación, De Vries les dio el nombre de mutaciones. El hecho significativo es la discontinuidad. Al físico le recuerda la teoría cuántica, según la cual no hay energías intermedias entre dos niveles energéticos contiguos. Podríamos llamar la teoría de la mutación, de forma figurada, la teoría cuántica de la Biología. Más adelante veremos que tal denominación es mucho más que figurativa. Las mutaciones se deben, de hecho, a saltos cuánticos en las moléculas del gen. Pero la teoría cuántica solo tenía dos años cuando de Vries publicó su teoría de la mutación, en el año 1902. No es, pues, extraño que se necesitase una generación más para descubrir la íntima relación entre ambas.
3.2. Las mutaciones se heredan perfectamente
Las mutaciones se heredan tan perfectamente como los caracteres inalterados originales. A modo de ejemplo, en la primera cosecha de cebada que veíamos antes, unas pocas espigas podían tener una longitud de barbas o aristas considerablemente alejada del nivel de variabilidad dado en la figura 1: digamos que están totalmente desprovistas de aristas. Podríamos representar una mutación de De Vries y en tal caso se reproducirían realmente, lo cual significa que todos los descendientes carecerían de aristas. Una mutación es, claramente, un cambio en el tesoro hereditario y debe atribuirse a algún cambio en la estructura material de la sustancia de la herencia. De hecho, la mayoría de los cruces experimentales de importancia, los cuales nos han revelado el mecanismo de la herencia, consisten en el análisis detallado de la descendencia obtenida al cruzar, de acuerdo con un plan preconcebido, individuos mutados (o, en muchos casos, múltiplemente mutados) con individuos no mutados o mutados de otra forma.
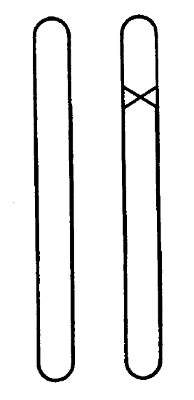
F IGURA 2. Mutante heterozigótico. La cruz señala el gen mutado.
Ademas, en virtud de su legítima transmisión, las mutaciones son un material apropiado sobre el cual puede operar la selección natural y producir las especies, según describió Darwin, eliminando los menos dotados y dejando sobrevivir a los mías aptos. Basta con sustituir en la teoría de Darwin «pequeñas variaciones accidentales» por «mutaciones» (de la misma forma que la teoría cuántica sustituye «transferencia continua de energía» por «salto cuántico»). Por lo demás, se requieren muy pocos cambios más en la teoría de Darwin, siempre que yo este interpretando correctamente el punto de vista de la mayoría de biólogos.
3.3. Localización. Recesividad y dominancia
Debemos revisar ahora otros hechos y nociones fundamentales referentes a las mutaciones, y de nuevo de forma algo dogmática, sin revelar directamente cómo se infieren, uno por uno, a partir de la evidencia experimental.
Podríamos esperar que una determinada mutación fuera causada por un cambio en una región determinada de uno de los cromosomas. Y así es. Es importante señalar que sabemos positivamente que se trata de un cambio en solo un cromosoma, pero no en el locus correspondiente del cromosoma homologo. La figura 2 indica esto esquemáticamente, representando la cruz el locus mutado. El hecho de que sólo se vea afectado un cromosoma se revela cuando el individuo mutado (llamado frecuentemente «mutante») se cruza con un individuo no mutado: exactamente la mitad de la descendencia presenta el carácter mutante y la otra mitad el carácter normal. Esto es lo esperable como consecuencia de la separación de los cromosomas en la meiosis del mutante, tal como se ve en la figura 3 de forma muy esquemática. En ella se ve un árbol genealógico que representa cada individuo (de tres generaciones consecutivas) simplemente por el par de cromosomas en cuestión. Tómese nota de que, si el mutante tuviera los dos cromosomas alterados, todos los hijos recibirían la misma herencia (mezclada), distinta de la de cualquiera de ambos padres.
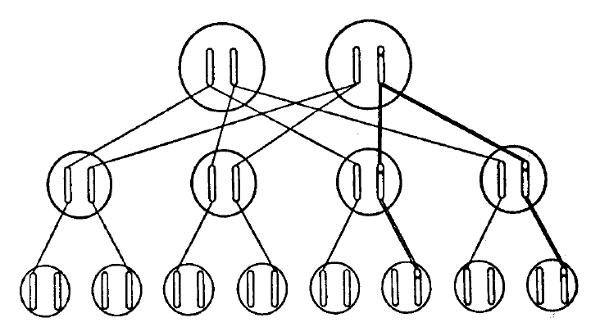
F IGURA 3. Herencia de una mutación. Las líneas rectas indican la transferencia de un cromosoma, las dobles, de un cromosoma mutado. Los cromosomas incorporados en la tercera generación proceden de las «paredes» de la segunda generación, que no se incluyen en el diagrama. Se supone que serón no relativos, ajenos a la mutación
Tamaño de fuente:
Intervalo:
Marcador:
Libros similares «¿Qué es la Vida?»
Mira libros similares a ¿Qué es la Vida?. Hemos seleccionado literatura similar en nombre y significado con la esperanza de proporcionar lectores con más opciones para encontrar obras nuevas, interesantes y aún no leídas.
Discusión, reseñas del libro ¿Qué es la Vida? y solo las opiniones de los lectores. Deja tus comentarios, escribe lo que piensas sobre la obra, su significado o los personajes principales. Especifica exactamente lo que te gustó y lo que no te gustó, y por qué crees que sí.